Genetisk variasjon og innavl hos norsk elg
Elgen er i dag en svært tallrik art og har en nær kontinuerlig utbredelse gjennom Skandinavia, det nordlige Eurasia og Nord-Amerika. Historiske kilder forteller imidlertid at det for 200 år siden kun eksisterte små og spredte bestander i Skandinavia og det nordlige Eurasia, men det er usikkert i hvilken grad disse utvekslet individer og gener. Inntil nylig visste vi lite både om elgens genetiske variasjon eller dens evne til å takle forandringer i miljøet. Høy genetisk variasjon kan for eksempel være avgjørende for at en art skal kunne takle klimaendringene som forventes av FNs klimapanel. For å undersøke hvorvidt genetisk variasjon har blitt tapt fra norske elgbestander, samt for å få en bedre oversikt over den genetiske bestandsstrukturen, analyserte vi derfor genetisk variasjon på tvers av hele den norske bestanden. Prosjektet ble igangsatt i 2007 og noen resultater har tidligere blitt presentert i Hjorteviltet (2009).
To genetisk forskjellige underbestander

Vevsprøver for genetiske analyser klippes vanligvis ut av elgens øre under merking, eller en lite muskelbit skjæres fra slaktet etter at elgen er skutt. Her er det primus motor Ole A. Davidsen på Vega som forestår operasjonen. Foto: Kari Bjørneraas.
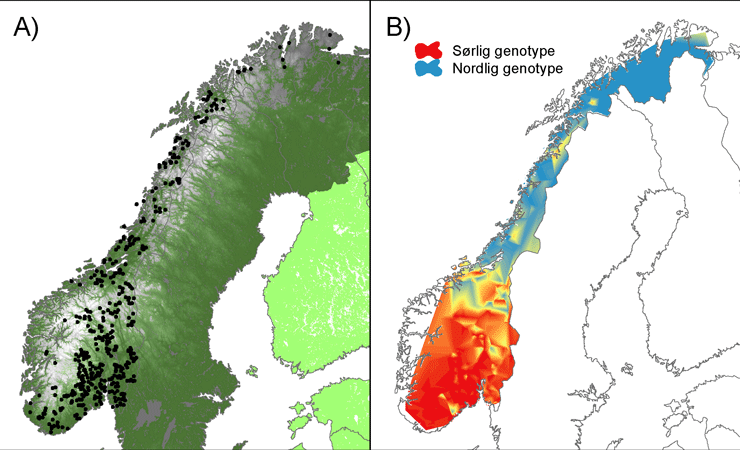
Undersøkelsen av genetisk variasjon i den norske elgbestanden viste at hver av 15 genetiske markører (mikrosatellitter) hadde en betydelig grad av variasjon, med 4 til 13 genetiske varianter (allel) per markør (snitt; 7,4). Den genetiske diversiteten varierte fra 0,30 til 0,79 mellom markørene (i snitt 0,66). Dette tyder på at den samlede genetiske variasjonen i den norske bestanden ikke er unormalt lav sammenlignet med andre hjortedyr. Det er derfor lite som tyder på at norsk elg gjennomgikk en genetisk flaskehals med tap av genetisk variasjon for 100 – 300 år siden. På den annen side viste de individbaserte analysene av genetisk likhet at den norske elgbestanden er delt opp i to genetisk forskjellige underbestander, en nordlig og en sørlig (Fig. 1). Utbredelsen av disse overlapper i et område i Sør-Trøndelag som ser ut til å ha liten utveksling av gener med både den nordlige og den sørlige bestanden. Dette tyder på at fjellområdene Dovre, Rondane, Forelhogna og Sylane, samt Trondheimsfjorden, utgjør betydelige barrierer for spredning. Interessant nok ser vi at elgen i Finnmark og den nordre delen av Troms også har vært påvirket av innvandring, antagelig fra Finland og/eller Russland.
Den genetiske strukturen i den norske elgbestanden er i samsvar med de historiske kildene som antyder at elgen var utryddet fra det meste av Norge i første halvdel av 1800-tallet, og kun var tilbake i deler av Hedmark og Trøndelag. I tillegg fantes et fåtall små bestander i Sverige, Finland og Russland. Dagens bestand kan i så fall være et resultat av ekspansjon og spredning fra en eller flere av disse kildebestandene. Alternativt kan todelingen stamme fra genetiske forskjeller som oppstod før bestandsnedgangen på 17- og 18-hundretallet. I nord, hvor elgen ifølge skriftlige kilder var helt utryddet, kan dagens bestand skyldes innvandring fra både midt-Norge, Sverige, Finland og Russland. Når vi beregnet nedarvingen til de observerte genetiske variantene (allel) bakover i tid fant vi imidlertid at dagens underbestander skyldes en genetisk bestandsoppdeling som er mer enn 1000 år gammel. Dette kan tyde på at dagens genetiske bestandsstruktur hovedsakelig skyldes at den norske elgen har fulgt innvandringsveier fra både nord og sør. Dette var mulig rett etter istiden da det var både en nordlig (som nå) og sørlig landforbindelse til kontinentet.
Et todelt innvandringsmønster med genetisk overlapp i Midt-Skandinavia er også beskrevet for bjørn, en del gnagere og flere plantearter. Bestander innen en art som isoleres over store geografiske avstander over lang tid utvikler ofte genetiske forskjeller som kommer til uttrykk i bestandsspesifikke tilpasninger og karaktertrekk. Dette kan også være tilfelle for den norske elgen som gjennomgående er større i nord enn sør. Dette har tidligere vært forklart med at elgen i nord og sør lever i ulike miljø med forskjellig temperatur og næringstilgang, men vi ville også undersøke hvorvidt genetisk opphav kan ha betydning.
I all hovedsak fant vi at elg fra den nordlige genetiske underbestanden hadde høyere slaktevekter enn elgen i sør, selv om vi tok hensyn til miljøforskjellene mellom områdene. Dette støtter hypotesen om at elgen har kolonisert Norge fra sør og nord, og at elgens karaktertrekk ikke kun skyldes dagens leveforhold, men også tilpasninger til ulike førhistoriske miljø.

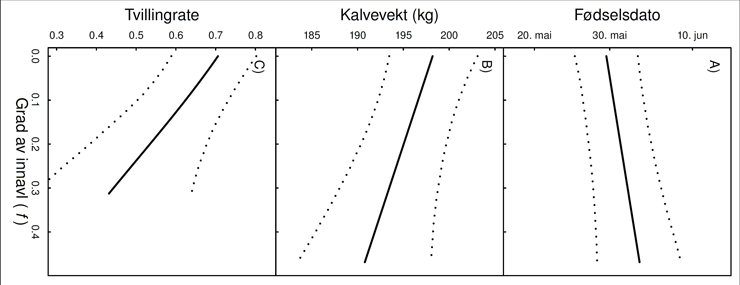
Figur 2. Graden av innavl (f) hos elgen på Vega i studieperioden 1985-2010. Høyere verdier antyder høyere innavlsgrad. Lavere verdier etter 1990 skyldes innflytelsen fra innvandrende individer.
Isolasjon og innavlseffekter på Vega
I tillegg til tap av genetisk variasjon og tilpasningsevne ved bestandsreduksjon, kan også innavl være et problem i små og isolerte bestander. Hos husdyr er det vist at innavl kan ha negative effekter på overlevelse, tilvekst og reproduksjon. Slike studier krever imidlertid god kunnskap om slektskapet mellom individene, og følgelig har vi få gode eksempler fra små og isolerte ville bestander. Ett godt eksempel er imidlertid elgbestanden på øya Vega i Nordland. Øya ble kolonisert av tre elg i 1985, og gjennom omfattende overvåking og forskning har vi siden fått en svært god oversikt over bestandens utvikling og biologi. Nylig benyttet vi også genetiske data fra disse elgene
til å utarbeide et nært komplett 25 års slektstre for bestanden, og med dette har vi nå et ideelt utgangspunkt for studier av innavl og innavlseffekter i en liten og isolert elgbestand.
Sammenlignet med den nordnorske fastlandsbestanden fant vi som ventet at elgen på Vega har betydelig færre genetiske varianter og lavere genetisk variasjon. Dette kan forklares med at få elg grunnla Vega-bestanden (3 individer), lav innvandring (20 individer) og en liten bestandsstørrelse siden kolonisering. Over halvparten av alle kalvene var koblet til en ku ut fra feltobservasjoner og blant disse ble de aller fleste (93 %) bekreftet å være mor-avkom av de genetiske slektskapsanalysene. Slektstreet ble også brukt til å regne ut graden av innavl hos hvert individ og viste at denne var relativt høy på Vega. Innavl uttrykker i hvilken grad genene et individ har mottatt fra moren og faren har lik avstamning (er i slekt). Dersom foreldrene er i slekt, kan skadelige enkeltmutasjoner fra begge havne hos avkommet, med potensielt svært negative konsekvenser.
Graden av innavl måles gjerne ved hjelp av innavlskoeffisienten. Denne kan variere fra null hos ikke-innavla dyr, og øker fra 0,06 ved paring mellom kusine og fetter, til 0,13 når halvsøsken parer seg, og til 0,25 når helsøsken parer seg. Graden av innavl kan også hope seg opp gjennom slektsledd. På Vega var tre fjerdedeler av individene innavla, og innavlsgraden økte over tid, men ble redusert i perioder med innvandring (Fig 3). I tillegg varierte innavlsgraden mye mellom innavla individer (0,02 – 0,47, snitt; 0,18). Det var imidlertid få individer fra parringer mellom svært nære slektninger (Av totalt 250 paringer, var kun 29 parringer mellom bror og søster eller foreldre og avkom).
Analysene viste at både fødselstidspunkt, kalvevekter og kuenes reproduksjon var relatert til graden av innavl (Fig. 3). Innavla individer ble født senere på våren og hadde en lavere kalvevekt, og innavla kyr fødte færre tvillinger. Kalvevekt var også relatert til fødselstidspunktet, mens tvillingrate var relatert til fødselstidspunkt og kalvevekt. Effekter av innavl forplanter seg således fra tidligere til seinere livsstadier. Analysene viste også at det var separate effekter av innavl på kalvevekt og tvillingrate, uavhengig av fødselstidspunktet.
Lignende innavlseffekter har blitt dokumentert hos enkelte andre ville hjortedyr og kan i verste fall forsterke de negative effektene av klimaendringer. I følge FN’s klimapanel vil vi i framtiden få en tidligere vår, noe som for elgens del kan medføre at kalvene blir født etter at plantene er som mest næringsrike på våren. Dersom innavl fører til ytterligere forsinkelser av kalvingen, kan dette ha store konsekvenser for hvor mye de vokser i løpet av sommeren. Små kalver blir vanligvis små voksne med gjennomgående lavere kalveproduksjon og overlevelse.
Figur 3. Endringer i kalvingstidspunkt, kalvevekt og tvillingrate med graden av innavl hos ulike elg på Vega.
Alt er relativt
Selv om innavl og innavlseffekter forekommer i norske elgbestander, tror vi det er liten grunn til å være bekymret for deres situasjon. Innavl er mest fremtredende i små bestander med liten innvandring fra andre bestander. I Norge er dette først og fremst tilfelle på øyer som ligger langt fra fastlandet, men kan muligens også opptre i fragmenterte deler av fastlandsbestanden. Av den grunn har vi også gjennomført genetisk analyser av elgbestandene på fastlandssiden av Vega, i et område med dype fjorder og høye fjell. Her var den genetiske variasjonen høyere, noe som antyder at utvekslingen av dyr mellom delbestander er tilstrekkelig stor til å unngå innavlseffekter som på Vega.
Vi tror heller ikke det er noen grunn til å bekymre seg for elgen på Vega. Til tross for innavlseffekter, er elgbestanden på Vega fortsatt i meget godt hold. Slaktevektene på kalv ligger jevnt mellom 70 og 80 kg, og nærmere 60 % av de reproduktive kyrne blir observert med tvillingkalver under jakta. Dette er egenskaper som bare de færreste bestandene i Sør-Norge kan framvise. Det heldige for Vega-elgen er at de lever i et miljø som har rikelig med høykvalitetsmat og med en relativt lang vekstsesong og snøfattig vintre. Dette kompenserer i stor grad for den kondisjonsreduksjonen som følger av at flere av individene er svært innavlede.
Sett under ett viser disse resultatene at elgens genetikk er et resultat av forskjellige prosesser på ulike nivå. Allerede da elgen innvandret rett etter siste istid hadde den sannsynligvis med seg forskjellig genetisk bagasje, avhengig av om den kom fra nord eller sør, og siden har varierende bestandsstørrelse og utbredelse påvirket den genetiske sammensetningen.
Elgens genetikk og tilpasningsevne er imidlertid gjenstand for kontinuerlig påvirkning og avhengig av påvirkningsfaktorenes retning og styrke vil dette kunne ha negative eller positive konsekvenser for elgen. En av årsakene til at dette genetikkprosjektet ble igangsatt, er bekymringen for at vår elgforvaltning også kan tappe elgbestandene for genetisk variasjon og tilpasningsevne. Hvorvidt disse bekymringene er berettiget eri øyeblikket gjenstand for nærmere analyser. Svaret, når det foreligger, vil forhåpentligvis danne grunnlaget for en ny artikkel i Hjorteviltet.
En takk rettes til: Jegere og forvaltere over det ganske land som har bidratt med vevsprøver fra elg, samt Norges forskningsråd, Miljødirektoratet og en rekke kommuner som har støttet prosjektet økonomisk.
LES MER:
Haanes, H., Røed, K.H., Solberg, E.J., Herfindal, I. og Sæther, B.-E. (2011) Genetic discontinuities in a continuously distributed and highly mobile ungulate, the Norwegian moose. Conservation Genetics 12(5): 1131–1143.
Haanes, H., Markussen, S., Herfindal, I., Røed, K.H., Solberg, E.J., Heim, M., Midthjell, L. Og Sæther, B.-E. (2013) Effects of inbreeding on fitness-related traits in a small isolated moose population. Ecology and Evolution 3(12): 4230–4242.
Herfindal, I., Haanes, H., Solberg, E.J., Røed, K.H., Høgda, K.A. og Sæther, B.-E. (2014) Moose body mass variation revisited: disentangling effects of environmental conditions and genetics. Oecologia 174(2): 447–458.
Solberg, E. J., Heim, M. Rolandsen, C. M., Sæther, B-E. & J. M. Arnemo 2011. Immobilisering og merking av elg på Vega, 1992-2010 – Konsekvenser for dyrevelferd, kondisjon, reproduksjon og overlevelse. – NINA Rapport 658. 30s. http://www.nina.no/archive/nina/PppBasePdf/rapport/2011/658.pdf