Fra vinter til sommerbeite – kan gi flere tvillingfødsler

Tidligere undersøkelser av elgen i Nord-Trøndelag har vist at det er stor variasjon i hvor store arealer dyrene bruker. Flere av elgene langs kysten har årsleveområder på noen få kvadratkilometer, mens andre vandrer kortere eller lengre distanser mellom sommer- og vinterleveområder. Data fra 144 GPS-merka elg ble benyttet for å undersøke hvorvidt trekkende elg hadde bedre tilgang på frisk og næringsrik vegetasjonen om våren og sommeren enn stasjonære elg, og om kalveproduksjonen og overlevelsen varierte mellom disse to gruppene.
Elgen trekker også oppover
Elgen er en delvis trekkende art. Det vilsi at noen oppholder seg i det samme området året rundt, mens andre trekker mellom separate sommer- og vinterområder. I tillegg har vi de som er en mellomting mellom stasjonær og trekkende. Det er lett å se for seg at de som trekker titalls av kilometer hver høst og vår kan kategoriseres som trekkelg. Men det er ikke bare de som vandrer langt som har ulike leveområder om sommeren og vinteren. Mange elg vandrer oppover i høyden om sommeren fra lavereliggende vinterområder, men ikke særlig langt i horisontal distanse. Også disse kan vi karakterisere som trekkende elg.
I gjennomsnitt tilbakelegger elg i Nord-Trøndelag en distanse på 17-18 km i løpet av vår- og sommermånedene og befinner seg omkring 100-150 meter høyere i terrenget på sommeren enn på vinteren (figur 1). Dersom vi kun ser på trekkende individer finner vi at vårtrekkets lengde hos disse er omkring 38 km i snitt for okser og 25 km for kyr.

Figur 1. Figuren viser hvor langt elgene i snitt har gått fra 15. april til de ulike datoene utover sommeren (akkumulert distanse). Data er fra 144 GPS-merka elg i Nord-Trøndelag i 2006–2010. Trekanter er okser, sirkler er elgkyr. Det er ikke skilt mellom stasjonære og trekkende individer.
Trekker kua helt til kalvinga?
Elgoksene går lengre enn kyrne. Det kom tydelig fram da vi så på elgenes GPS-posisjoner. I tillegg beveger oksene seg flere høydemeter oppover i landskapet (figur 1). Om våren og tidlig på sommeren har imidlertid kyr og okser relativt likt bevegelsesmønster, men i overgangen mai/juni endrer dette seg. På dette tidspunktet avtar den tilbakelagte distansen for kyrne seg betraktelig. Dette sammenfaller med kalvingstidspunktet: Gjennomsnittskua i Nord-Trøndelag kalver i starten av juni. Det kan altså virke som om kua enten når fram til sitt sommerområde tidligere enn oksene, eller at kalvene begrenser kuas mulighet til å forflytte seg i tiden rundt og like etter fødsel slik at hun forblir i det området hun kalver i.
Hvorfor trekke på våren?
Elgen starter gjerne å trekke mot lavereliggende vinterområder når snøen ankommer. Men hva gjør at elgen vandrer ut av disse områdene når våren kommer?
Det er mange hypoteser knyttet til dette spørsmålet. Studier av hjortevilt andre steder har vist at dyrene forflytter seg mellom sommer- og vinterområder for å unngå rovdyr eller parasitter. Høy tetthet og konkurranse med artsfrender er en annen årsak til at hjortevilt trekker mellom sesongområder. Det som mange forbinder med vårtrekket, er imidlertid dyr som følger endringer i vegetasjonen. Dette er for eksempel tydelig for flere arter på de store, åpne savannelandskap i Afrika, hvor romlig variasjon i nedbør skaper store forskjeller i hvor beitedyrene finner frisk vegetasjon.
Planteutviklingen – kan den utnyttes til elgens fordel?
Norge har langt i fra noe afrikansk savannelandskap, men noen likheter kan vi likevel påberope oss. Snøsmeltinga om våren skjer til ulik tid i innland og ved kysten og varierer med høyde over havet. Dette fører til at vegetasjonen starter å spire til ulik tid i ulike områder, noe elgen potensielt kan utnytte. Nyspirte planter har høy næringsverdi og er lette å fordøye for elgen. Individer som har tilgang til mye frisk vegetasjon kan derfor dra fordel av dette, for eksempel gjennom økt vekst eller reproduksjon.
I innlandet eller høyereliggende områder foregår snøsmelting og spiring av planter på et senere tidspunkt enn i lavlandet og ved kysten. Ved først å utnytte frisk vegetasjon i de sistnevnte områdene, for så å vandre i bakkant av snøsmeltinga innover i landet eller oppover i høyden, kan elgen forlenge tiden den har tilgang til mat av høy kvalitet.
Elg som går langt og høyt får bedre tilgang på kvalitetsmat
Vi undersøkte om elgens bevegelsesmønster om våren faktisk var relatert til vegetasjonsutviklingen. I Nord-Trøndelag er plantenes vekstsesong fra rundt mai til september, men varierer fra kyst til innland og med høyden over havet. Disser landskapsgradientene skaper altså en asynkron vekstsesong som elgen teoretisk sett kan utnytte. For å undersøke om den faktisk også gjør dette, undersøkte vi hvordan GPS-merka okser og kyr beveget seg i landskapet i forhold til plantenes utvikling.
Resultatene fra våre undersøkelser viste at elgenes bevegelser om våren og sommeren var relatert til plantenes utvikling. Det vil si at elgene på et gitt tidspunkt oppholdte seg i områder hvor vegetasjonen var friskere og av bedre kvalitet enn i områdene den brukte ellers i vekstsesongen. Hvor tett elgene fulgte vegetasjonsutviklingen varierte imidlertid mellom individer. Særlig de elgene som gikk langt i løpet av våren og sommeren, og de som bevegde seg oppover i terrenget, fikk en lengre periode med tilgang til frisk vegetasjon. Elgokser utnyttet dessuten gradienten i plateutviklingen bedre enn kyr, kanskje fordi oksene generelt går både lengre og høyere i terrenget enn hunndyra. Felles for begge kjønn var imidlertid at individer som oppholder seg mye nær dyrka mark ikke fulgte gradienten i planteutviklinga like tett som andre individer. Dette kan blant annet være fordi dyrka mark er en god matkilde i seg selv og elgene vil derfor ikke ha det samme behovet for å utnytte vegetasjonsendringene i utmarka.
For hjort er det gjort tilsvarende undersøkelser i Norge, og her fant forskerne ut at hjorten «hopper» fra en tidlig vår i lavlandet til en ny og senere vår i høyden. Hjorten bruker så kort tid på å trekke mellom sommer- og vinterområdene at vi ikke kan si at den følger planteutviklinga. Elgen i Nord-Trøndelag bruker betraktelig lenger tid på trekkruta om våren, i gjennomsnitt ca. 13 dager, men igjen er det stor variasjon i antallet dager elgene trekker. Våre GPS-merka elg brukte mellom 1 – 60 dager mellom vinter- og sommerområdet. Vi kan ikke med sikkerhet si at elgen følger i kanten på snøsmeltinga da vegetasjonsdataene har et tidsintervall på 16 dager. Det vi med rimelig sikkerhet kan si, er imidlertid at de elgene som trekker får bedre tilgang på friske og lettfordøyelige planter enn de ville gjort om de forble året rundti vinterområdene.

Elg som trekker får bedre tilgang til kvalitetsmat. Foto Line Venn
Trekkende elg har oftere tvillingkalver
Nå vet vi at nordtrønderske elg trekker langt eller beveger seg oppover i terrenget får tilgang til planter av høy kvalitet over lengre tid. Spørsmålet som da dukker opp er om dette hadde noen påvirkning på dyrenes kondisjon og fruktbarhet? For å undersøke dette sammenlignet vi tvillingproduksjonen hos 38 trekkende og 46 stasjonære elgkyr der antallet kalver (en eller to) var kjent.
Undersøkelsene viste at elgkyrne som trakk mellom sommer- og vinterleveområder hadde 1,7–2,5 ganger større sannsynlighet for å ha tvillinger enn stasjonære kyr, og dette synes å være uavhengig av alderen på dyra (figur 2). Motsatt var det ingen ting som tydet på at trekkende og stasjonære elgkyr var forskjellige med hensyn til drektighetsrate.
Vi undersøkte også om sannsynligheten for å overleve var ulik mellom trekkende og stasjonære elgkyr, og tilsvarende om kalvene til trekkende og stasjonære elgkyr viste forskjellig dødelighet. Det var en svak tendens til at trekkende elgkyr mistet en lavere andel kalv enn stasjonære elgkyr i løpet av vinteren, men denne forskjellen var ikke statistisk sikker.
Med andre ord er det god grunn til å tro at trekkende individer produserer langt flere kalver enn stasjonære individer og at en minst like stor andel trekkende kyr og deres kalver overlever vinteren. Ved å sammenligne tettheten av beitetrær i områder med trekkende og stasjonære individer, ser vi dessuten at trekkende elg har mer mat tilgjengelig per individ i løpet av sommersesongen. Det store spørsmålet er derfor hvorfor ikke flere elgkyr utnytter muligheten til å trekke fra vinterområder til eksklusive sommerområder? Dette er noe vi så langt ikke har noe godt svar på, men det hindrer oss ikke fra å spekulere.
En mulighet er at dagens store forskjeller i kalveproduksjon mellom trekkende og stasjonære mødre er et relativt nytt fenomen som skyldes et varmere klima. Dette har gitt en lengre vekstsesong i mange deler av landet, noe som kan ha gavnet trekkende elgkyr, som kalver i innlandet eller høyere til fjells, mer enn det gavnet stasjonære elgkyr.
For eksempel kan det nå være et bedre sammenfall mellom plantenes utvikling og kalvingstiden for kyr i høyreliggende områder enn hva det var tidligere. God tilgang til næringsrik føde er fordelaktig for kua i den siste delen av drektigheten og den første delen av kalvens liv. Til sammenligning er det mulig at kyr i lavereliggende områder nå opplever et mindre optimalt sammenfall mellom plantenes utvikling og kalvingsdato, med dertil negative effekter på kalveproduksjonen.
Dersom klimaendringene faktisk skaper slike forskjeller i kalveproduksjonen er det grunn til å tro at andelen trekkende elg i Nord-Trøndelag vil øke i årene som kommer. En forutsetning er dog at kalvene til trekkende individer faktisk adopterer den samme strategien som mora. Selv om det er grunn til å tro at det eksisterer en slik sammenheng, er det ikke kjent i hvor stor grad dette er tilfelle. Dersom trekkatferden kun i liten grad arves fra foreldre til avkom (kulturelt eller genetisk), kan det ta lang tid før vi ser slike endringer.
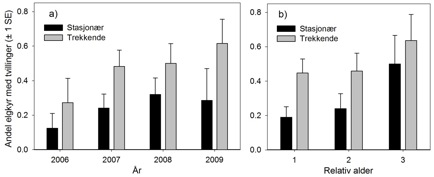
Figur 2. a) Årlig tvillingrate (± 1 standardfeil) for stasjonære (svarte stolper) og trekkende (grå stolper) elgkyr. b) Årlig tvillingrate (± 1 standardfeil) i forhold til kyrenes relative alder, dvs. alder siden merking (merkeår = relativ alder 1).
Konklusjon
Våre undersøkelser tyder på at det er fordelaktig å være trekkelg i vårt studieområde. Elg som går langt eller klatrer høyt om våren og sommeren får bedre tilgang til mat av god kvalitet over tid. Ser vi bort fra jordbruksområdene langs kysten, har også elg som oppholder seg i høyden bedre mattilgang per dyr i sommerhalvåret enn elg lavere i terrenget. Disse forskjellene i mattilbud sammenfaller med at trekkende elg også har vesentlig større sannsynlighet for å produsere tvillinger enn stasjonære elg. Dersom disse forskjellene forblir på det samme nivået i årene som kommer er det grunn til å tro at andelen trekkelg i Nord-Trøndelag vil øke, men dette forutsetter at kalvene faktisk følger i moras fotspor. Trekkende elg oppfattes av mange som et problem fordi inntekter (jakt) og kostnader (skog- og trafikkskader) ofte faller på forskjellige grunneiere og kommuner. Samtidig ser vi her at trekkmuligheten også kan gi elgen bedre muligheter til å utnytte de forskjellene i matkvalitet som eksisterer i norsk natur.
Litteratur:
Christer M. Rolandsen. The ecological significance of space use and movement patterns of moose in a variable environment. PhD-avhandling. NTNU 2012.